






Ботулинический токсин (БТ) нашел свое применение в урологии при детрузорной гиперактивности нейрогенной и идиопатической этиологии у пациентов, невосприимчивых к медикаментозной терапии холинолитиками [1, 2].Нарушение афферентной и эфферентной иннервации мочевого пузыря лежит в основе детрузорной гиперактивности [3-5], а анальгезирующий эффект БТ связывают с его влиянием на нервную афферентацию [6]. Регуляция функции мочевого пузыря осуществляется центральной нервной системой посредством координированных симпатических и парасимпатических влияний. Однако выявить значение разных отделов вегетативной иннервации для его функции является сложной задачей. В нашей предыдущей работе использована новая импедансная техника для изучения состояния кровообращения и регуляторных процессов, осуществляемых вегетативной нервной системой, в разные фазы активности нормального мочевого пузыря крыс и после моделирования частичной инфравезикальной обструкции [7].
На основании этих исследований высказана гипотеза о том, что пик Майера и респираторный пик в спектре импеданса мочевого пузыря могут характеризовать его нейрогенную (соответственно, симпатическую и парасимпатическую) активность. Основанием для этого послужили наблюдения о противоположных изменениях этих пиков в разные фазы функциональной активности нормального мочевого пузыря, - рост пика Майера во время накопительной фазы при одновременном снижении респираторного пика и напротив, увеличение респираторного пика при уменьшении пика Майера во время эвакуаторной фазы. Такие изменения, по нашему мнению, отражают реципрокную регуляцию функции мочеиспускания симпатическим и парасимпатическим отделами вегетативной нервной системы. Кроме того, в мочевых пузырях крыс после моделирования инфравезикальной обструкции были выявлены нарушения этих регуляторных процессов.
Настоящее исследование посвящено исследованию симпато-парасимпатической активности и уродинамики нормального и обструктивного мочевого пузыря после инъекций БТ в детрузор.
МАТЕРИАЛ И МЕТОДЫ
Исследования выполнены на анестезированных крысах (эфирно-хлоралозный наркоз) в условиях открытого оперативного доступа к мочевому пузырю. Эксперименты проведены на 15 интактных крысах, а также 11 крысах с моделированной частичной инфравезикальной обструкцией (ИВО), вызванной перевязкой уретры на предварительно введенном катетере за 1 месяц до исследования. Кроме того, 5 интактным крысам и 6 крысам через 1 месяц после моделирования ИВО инъецировали ботулинический токсин (БТ) «Лантокс» в стенку мочевого пузыря (3 ед. на крысу); обследование производили через 7-10 дней после инъекций. Соответственно, крыс разделили на 4 группы: контрольную - «контр» (n = 10), «ИВО» (n = 5), «контр+БТ» (n = 5) и «ИВО+БТ» (n = 6).
Детрузорное давление (Pdet) регистрировали в условиях открытой брюшной стенки электроманометром ТА-2 (производство НПФ «Биола», Россия) через цистостомический катетер, установленный в дно пузыря. Посредством этого катетера осуществляли инфузионную цистометрию роликовым насосом производительностью 0,09 и 0,9 мл/мин. при сохранении самопроизвольного мочеиспускания по интактному мочеиспускательному каналу. В отдельных экспериментах в мочевой пузырь вводили атропин в составе инфузионного физиологического раствора (0,2 мг/л). Для отведения реограммы (импеданса) мочевого пузыря использованы два проволочных кольцевидных хлорсеребряных электрода, фиксируемых наружно в области тела пузыря и ближе к его шейке. Одновременную регистрацию показателей давления, базового (полного) импеданса и его переменной составляющей осуществляли оригинальным программно-аппаратным комплексом (НПФ «Биола», Россия), включающим высокочувствительный и малошумящий импедансметр с разрешением 250 микроом по каналу переменной составляющей, 4-канальный преобразователь ППШ-04 и ЭВМ. Для анализа вариаций импеданса мочевого пузыря использовали Фурье преобразование в диапазоне частот от 0,05 Гц до 15,0 Гц. Частота дискретизации составляла 160 Гц. Для анализа выбирали фрагменты длиной 12,8 или 25,6 секунд, которые соответствовали определенному функциональному состоянию мочевого пузыря. Измерения проводили в разные периоды функциональной активности мочевого пузыря: исходное состояние («покой»), во время наполнения физиологическим раствором (инфузионная цистометрия), а также после окончания инфузии. Длительность записей составляла 4-минуты в каждом состоянии. Результаты обрабатывались статистически с использованием критерия Стьюдента.
Таблица 1. Функциональные показатели мочевого пузыря у крыс разных групп, зарегистрированные в состоянии покоя
| Группы крыс | Детрузорное давление (cм вод. ст.) | Мощность основных спектральных пиков импедансограммы мочевого пузыря (10-3Ом2) | |
| пик Майера | респираторный пик | ||
| Контр | 4,64 ± 1,40 | 13,97 ± 0,3 | 15,81 ± 0,4 |
| Контр + БТ | 2,50 ± 1,30 | 7,30 ± 0,6 | 2,08 ± 0,4 |
| ИВО | 8,23 ± 2,65 | 20,89 ± 0,6 | 21,27 ± 0,8 |
| ИВО + БТ | 4,18 ± 2,52 | 2,58 ± 0,12 | 1,34 ± 0,1 |
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
После инъекций БТ в стенку мочевого пузыря выявлены изменения функционального состояния детрузора и его нейрогенной регуляции у крыс с нормальным и обструктивным мочевым пузырем (таблица 1).
Детрузорное давление у крыс с обструктивным мочевым пузырем было выше на 77% по сравнению с контрольной группой крыс. В состоянии покоя диапазон его изменений составлял 5-15 см вод. ст., по сравнению с 1-3 см вод. ст. в контрольной группе крыс с неизмененными мочевыми пузырями. При этом средняя мощность пика Майера у крыс с ИВО была больше на 50%, а мощность респираторного пика на 34% больше, чем в контрольной группе крыс. Таким образом, формирование ИВО ведет к активизации обоих звеньев вегетативной регуляции функции мочевого пузыря, что может быть связано с перестройкой его функциональной активности.
После инъекций БТ наблюдали более низкие, чем без БТ, значения внутрипузырного давления и мощности обоих пиков, характеризующих нейрогенную активность мочевого пузыря. Так, детрузорное давление было ниже после инъекционного введения БТ в стенку МП крыс с нормальным мочевым пузырем на 2,14 ± 0,55 см вод.ст. (-46%, р < 0,01) и крыс с моделированной обструкцией мочевого пузыря на 4,05 ±1,96 см вод. ст. (-49%, 0,1 < p >0,05) при сравнении этих показателей с соответствующими им группами крыс «контр» и «ИВО». При этом пик Майера и респираторный пик, отражающие нейрогенную активность мочевого пузыря, также были снижены после введения БТ у крыс с ИВО (-88% и -94% соответственно), и у крыс с прежде нормальными мочевыми пузырями (-48% и -83% соответственно). Полученные результаты объяснимы с позиций механизма действия БТ, который блокирует высвобождение ацетилхолина из холинергических нервных синапсов, в результате чего происходит химическая денервация мочевого пузыря и угнетение его нейрогенной активности.
Во время инфузионной нагрузки рост детрузорного давления сопровождался определенными изменениями спектральных характеристик импеданса мочевого пузыря в разных группах экспериментальных крыс, которым инъецировали токсин (рисунок 1).
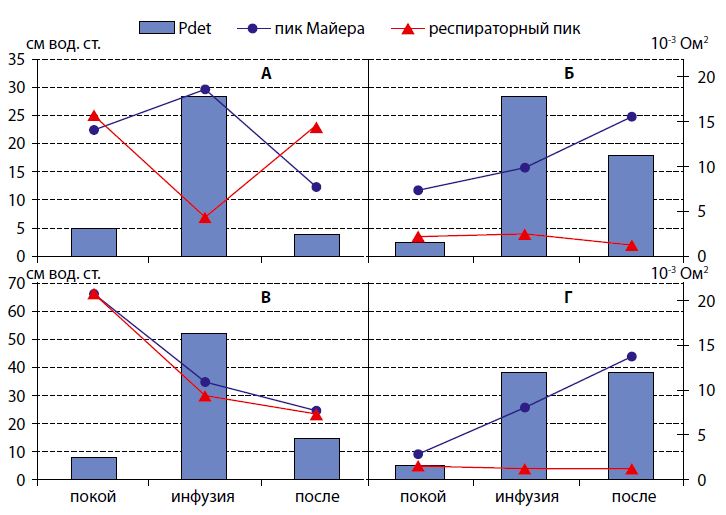 |
Рисунок 1. Изменение внутрипузырного давления (Pdet) и спектральных пиков импеданса мочевого пузыря, характеризующих нейрогенную активность, во время инфузионной цистометрии у крыс А – контрольной, Б – «контр + БТ», В – «ИВО», Г – «ИВО+БТ» групп. (Столбиками обозначены величины детрузорного давления в см вод. ст., а линейные диаграммы представляют динамику мощности пика Майера (кружочки) и респираторного пика (треугольники) в 10-3Ом2) |
В предыдущей работе было показано, что при наполнении мочевого пузыря у крыс контрольной группы (рисунок 1A) внутрипузырное давление возрастало почти в 6 раз, при этом мощность волны Майера увеличивалась (+32%). После прекращения инфузии раствора в пузырь и снижения детрузорного давления происходило уменьшение пика Майера на 75% от исходного уровня, наблюдаемого до инфузии. Изменения респираторного пика во время инфузионной цистометрии имели противоположный характер: его мощность уменьшалась в начале инфузии раствора в пузырь (-73%) и увеличивалась до 92% от исходного уровня в конце процедуры, когда происходило выделение жидкости по уретре.
У крыс с ИВО (рисунок 1В) наблюдали более выраженный прирост детрузорного давления во время инфузионной цистометрии, чем в контрольной группе, - средний прирост детрузорного давления в ответ на нагрузку превышал прирост давления в контрольной группе крыс на 88%, и его величина оставалась повышенной после прекращения инфузионной цистометрии на 81% по отношению к исходному уровню, зарегистрированному до начала инфузии раствора в пузырь. Однако, несмотря на высокие подъемы давления в пузыре во время его наполнения у крыс с ИВО, показатели импедансного спектра мочевого пузыря, отражающие нейрогенную активность, снижались во время и после инфузионной нагрузки. Мощность майеровского и респираторного пиков уменьшилась в среднем почти в 2 раза (-48% и -55% соответственно) во время инфузионной нагрузки в сравнении со значением до ее начала, и оставалась сниженной на 63-66% после инфузии. Следовательно, в динамике исследуемых показателей во время инфузионной цистометрии у крыс с ИВО не выявлено противоположных изменений волны Майера и респираторного пика, как в неизмененных пузырях контрольной группы, которые мы объясняем реципрокными связями симпатического и парасимпатического отдела вегетативной иннервации мочевого пузыря. В нормальных пузырях показатель отношения мощностей пика Майера и респираторного, отражающий симпатовагусный баланс, уменьшался от 4,3 до 0,5 при переходе от накопительной к эвакуаторной фазе, а в обструктивных пузырях этот показатель практически не менялся, составляя, соответственно 1,1 и 1,0. Эти результаты демонстрируют нарушения нейрогенной регуляции функции пузыря при ИВО, осуществляемых вегетативной нервной системой.
После инъекций БТ в стенку пузыря (рисунки 1Б и 1Г) наблюдалась иная динамика детрузорного давления и показателей импедансного спектра. В обеих группах экспериментальных крыс, которым инъецировали токсин, при неравнозначных подъемах детрузорного давления в ответ на одинаковую инфузионную нагрузку изменения спектральных характеристик импеданса мочевого пузыря были приблизительно одинаковые. Мощность волны Майера у крыс, которым токсин вводили в стенку прежде нормального мочевого пузыря (Б), увеличивалась при подъеме детрузорного давления до 9,8 ± 0,1 10-30м2 во время инфузионной нагрузки и достигала максимального значения 15,4 ± 0,8 10-30м2 по окончанию инфузии. У крыс, которым токсин вводили в обструктивный мочевой пузырь (Г), мощность волны Майера возрастала до 8,1 ± 0,1 10-30м2 во время наполнения мочевого пузыря и достигала максимально 13,8 ± 0,7 10-30м2. Мощность респираторного пика в импедансном спектре была низкой в обеих группах (диапазон 0,4-1,3 и 0,1-2,5 10-30м2 соответственно, в группах «контр + БТ» и «ИВО + БТ»). Таким образом, после введения БТ наполнение мочевого пузыря, которое должно стимулировать мочеиспускание, не сопровождалось усилением парасимпатической активности, свидетельствуя о блокаде этого нейрогенного звена регуляции.
Полученные результаты свидетельствуют об угнетении холинергического механизма регуляции его функции после действия токсина и активизации симпатической активности, оцениваемой по возрастанию пика Майера в абсолютных и в относительных значениях. Так, расчетный показатель отношения мощностей пика Майера и респираторного пика в импедансном спектре мочевого пузыря показывал смещение симпатовагусного баланса в сторону преобладания симпатической активности, причем этот сдвиг был более выражен у крыс, получавших инъекции БТ, с ИВО, чем с нормальными пузырями. Этот показатель был низким (3,8) в группе «контр + БТ» в фазу накопления, но возрастал до 12,8 во время эвакуаторной фазы. Тогда как в группе «ИВО + БТ» этот показатель составлял 8,1 во время резервуарной фазы и увеличивался лишь до 9,2 при эвакуа- торной фазе. С изменениями симпатического и парасимпатического звеньев вегетативной регуляции мочевого пузыря может быть связано наблюдаемое высокое детрузорное давление после прекращения инфузии в обеих группах крыс, получавших инъекции БТ. Также, более высокие значения давления в пузыре в группе «ИВО + БТ», по сравнению с группой контрольных крыс, получавших токсин, могут быть обусловлены изменениями симпато-вагусного баланса.
Таблица 2. Детрузорное давление у экспериментальных крыс при инфузионной цистометрии
| Время регистрации | Детрузорное давление (см вод. ст.) в группах крыс | |
| Контр + БТ | ИВО + БТ | |
| покой | 2,50 ± 1,30 | 4,18 ± 2,52 |
| инфузия 0,09 мл/мин. | 28,56 ± 6,52 | 37,63 ± 7,36 |
| инфузия 0,9 мл/мин. | 42,45 ± 15,89 | 67,08 ± 11,18 |
| после инфузии | 17,83 ± 9,90 | 38,00 ± 7,98 |
В таблице 2 представлены результаты цистометрических исследований у крыс, получавших инъекции БТ, при умеренном (0,09 мл/ мин.) и высоком (0,9 мл/мин.) режимах инфузионной нагрузки.
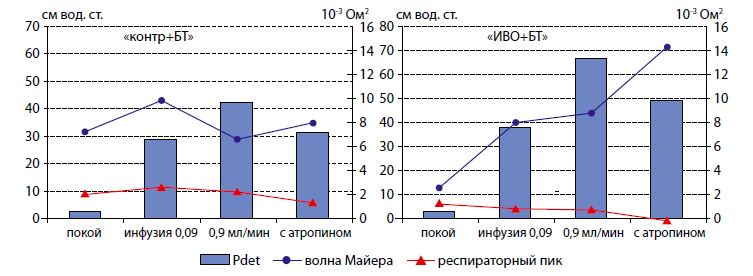 |
| Рисунок 2. Динамика детрузорного давления и пиков импедансного спектра при добавлении атропина в инфузионный раствор (обозначения – как на рисунке 1) |
Как показывают результаты измерений, давление в обструктивных мочевых пузырях после инъекции токсина превышало таковое в мочевых пузырях крыс «контр + БТ» группы. Детрузорное давление в пузырях у крыс группы «ИВО + БТ» в покое было выше на 1,7; 9,1; 24,6 и 20,2 см вод. ст. (на 67; 32; 58 и 113%) соответственно, при низком; высоком режимах цистометрии и по окончании инфузии физиологического раствора в пузырь, по сравнению с давлением у крыс группы «контр + БТ». Следовательно, в группе крыс с обструктивным мочевым пузырем этот уродинамический показатель в меньшей степени, чем у крыс с прежде нормальным мочевым пузырем, подвержен действию токсина.
Такие различия могут быть обусловлены разными механизмами формирования тонуса детрузора после формирования инфравезикальной обструкции. С одной стороны, в стенке обструктивного мочевого пузыря происходят структурные перестройки, приводящие к снижению его эластичности, что проявляется высокими значениями давления в пузыре при растяжении его стенки большим объемом инфузируемого раствора. С другой стороны, известна особая роль уротелия и его аутокринные и паракринные эффекты, обусловленные высвобождаемыми при растяжении стенки пузыря биологически активными пептидами и медиаторами нервного возбуждения [3, 8-11]. Эти регуляторные факторы могут влиять на функциональные изменения тонуса мочевого пузыря, обеспечиваемого собственно мышечными клетками детрузора и, следовательно, на показатель детрузорного давления.
Снижение детрузорного давления после инъекции токсина и угнетение мощности показателей спектра мочевого пузыря, характеризующих его нейрогенную активность, подтверждают факт химической денервации детрузора. Блокирование токсином высвобождаемого из нервных окончаний ацетилхолина способствует снижению миогенного тонуса, в результате чего происходит снижение давления в пузыре и увеличение его объема. Наблюдаемое более высокое детрузорное давление в разные периоды инфузионной цистометрии в группе крыс с обструктивным мочевым пузырем, в сравнении с нормальным пузырем, несмотря на введение токсина, указывает на возможное наличие структурных изменений в его стенке, которые с трудом могут поддаваться медикаментозному лечению. В то же время, выявление функциональных нарушений регуляции тонуса обструктивного детрузора может быть полезным для диагностики, т.к. их корректировка более вероятна. Проведенные исследования показали, что нарушения функциональной регуляции мочевого пузыря происходят после моделирования инфравезикальной обструкции. Эти регуляторные изменения в свою очередь, наряду со структурной перестройкой детрузора, могут быть причиной наблюдаемых более высоких значений давления в обструктивном мочевом пузыре, по сравнению с таковыми в нормальном пузыре, после введения токсина.
Для проверки роли холинорецепторного механизма регуляции функции мочевого пузыря после действия БТ в ряде экспериментов выполняли внутрипузырное введение атропина в составе инфузионного раствора. Атропин вызывал снижение детрузорного давления во время инфузии в режиме 0,9 мл/мин. у крыс «контр + БТ» с 42 до 31 см вод. ст. и с 67,1 до 49,0 см вод. ст. в группе крыс «ИВО + БТ» (рисунок 2).
При этом выявлены интересные особенности динамики спектральных характеристик нейрогенной активности. Атропин вызывал усиление мощности пика Майера и угнетение мощности респираторного пика не только в контрольных сериях («контр»,«ИВО»), но и в опытах с введением БТ, когда холинорецепторы должны быть инактивированы (рисунок 3). Зарегистрировано увеличение пика Майера на 42% и 85% в группах «контр» и «ИВО», а у крыс, получавших инъекции БТ, прирост был менее выражен и составлял 21% и 62% в группах «контр + БТ» и «ИВО + БТ» соответственно. После внутрипузырного введения атропина мощность респираторного пика уменьшалась на 17% и 58% в группах «контр» и «ИВО», а у крыс, получавших инъекции БТ, изменения были более выражены, составляя в группах «контр + БТ» и «ИВО + БТ» 41% и 90% соответственно.
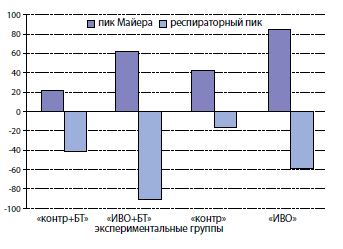 |
Рисунок 3. Относительные изменения пиков импедансного спектра мочевого пузыря при добавлении в инфузионный раствор атропина (0,2 мг/л). Изменения пиков импедансного спектра мочевого пузыря после введения в инфузионный раствор атропина (0,2 мг/л) представлены в % от их величин, зарегистрированных во время инфузионной цистотетрии с объемной скоростью 0,9 мл/мин физиологическим раствором, не содержащим атропин |
Наблюдаемая динамика показателей нейрогенной активности мочевого пузыря после инфузии с атропином, проявляющаяся уменьшенными реакциями пика Майера у ботулинизированных животных по сравнению с крысами, не получавшими токсин, объясняется угнетением нейрогенной активности после действия токсина, а увеличение реакций респираторного пика на атропин указывает на дополнительные источники медиаторов холинергических рецепторов. Блокада холинорецепторов уротелия растянутого мочевого пузыря инфузируемым физиологическим раствором с атропином вызывала изменения респираторного пика у крыс после моделирования ИВО, которые были более выраженными, чем у контрольных крыс. Это, вероятно, обусловлено особенностями автономной регуляции обструктивного мочевого пузыря и возросшей ролью ненейрогенного ацетилхолина, высвобождаемого уротелием, который считается особым нейромышечным синцитием. Роль этого медиатора и других, действие которых опосредуется холинорецепторами уротелия, возрастает при ИВО. Согласно нашим проведенным исследованиям можно предположить, что БТ блокирует реакции, опосредуемые холинергическими медиаторами, высвобождаемыми из нервных окончаний, однако мало влияет на функцию пузыря, опосредуемую ненейрогенными холинергическими медиаторами.
Наблюдаемое уменьшение реактивности пика Майера в группах крыс, которым выполняли инъекции БТ, по сравнению с соответствующими группами контрольных крыс, объясняется с позиций механизма действия токсина, вызывающего химическую денервацию органа и, следовательно, угнетение его нейрогенной активности. Тенденция к усилению отрицательных изменений респираторного пика при инфузионном введении атропина указывает на возросшую роль стимуляторов холинорецепторов ненейрогенного происхождения в мочевых пузырях крыс с ИВО, и получавших инъекции токсина.
ЗАКЛЮЧЕНИЕ
Проведенные экспериментальные исследования показали возможность оценки нейрогенной активности мочевого пузыря одновременно с детрузорным давлением для характеристики участия симпатического и парасимпатического компонентов вегетативной нервной системы в регуляции его резервуарной и эвакуаторной функций. Инъекционное введение ботулинического токсина в стенку пузыря вызывало угнетение обоих пиков импедансного спектра, при этом происходило существенное увеличение отношения пика Майера к респираторному пику, что свидетельствует об изменении вегетативного баланса в ботулинизированном мочевом пузыре в пользу симпатической нервной активности. Изменения вегетативной регуляции мочевого пузыря при ИВО, связанные с возросшей ролью ненейрогенных регуляторов функции детрузора, могут быть причиной повышенного тонуса детрузора, несмотря на действие БТ.
Ключевые слова: мочевой пузырь, вегетативная иннервация, инфравезикальная обструкция, импедансная цистометрия, ботулинотоксин.
Keywords: bladder, autonomic innervations, infravesical obstruction, impedance cystometry, botulinotoxini.
Литература
- Dmochowski R., Chaple C., Nittiv V.W. Efficasy and safety of onabotulinumtoxin A for idiopathic overactive bladder: a doubleblind, placebo controlled, randomized, dose ranging trial // J. Urol. 2010. Vol. 184, № 6. Р. 2416-2422.
- Smith C.P., Nishiguchi J., O’Leary M., Yoshimura N., Chancellor M.B. Single-institution experience in 110 patients with botulinum toxin A injection into bladder or urethra // Urol. 2005. Vol. 65, № 1. Р. 37-41.
- Apostolidis A., Dasgupta P., Fowlera C.J. Proposed Mechanism for the Efficacy of Injected Botulinum Toxin in the Treatment of Human Detrusor Overactivity // European Urology. 2006. Vol. 49, № 4. P 644-650.
- Nishijima S., Sugaya K., Kadekawa K., Naka H., Miyazato M. Comparison of the effect of anti-muscarinic agents on bladder activity, urinary ATP level, and autonomic nervous system in rats // Biomedical Research. 2009. Vol. 30, № 2. Р.107-112.
- Lawrence G.W., Aoki K.R., Dolly J.O. Excitatory Cholinergic and Purinergic Signaling in Bladder Are Equally Susceptible to Botulinum Neurotoxin A Consistent with CoRelease of Transmitters from Efferent Fibers// J Pharmacol Exp Ther. 2010. Vol. 334. Р. 1080-1086.
- Aoki K.R. Review of a proposed mechanism for the antinociceptive action of botulinum toxin type A // Neurotoxicology. 2005. Vol. 26, № 5. Р. 785-793.
- Мудрая И.С., Ибрагимов А.Р., Кирпатовский В.И., Ревенко С.В., Нестеров А.В., Гаврилов И.Ю. Оценка функционального состояния мочевого пузыря крыс методом Фурьеимпедансной цистометрии. Экспериментальная и клиническая урология. 2010. № 3. С. 21-26.
- Mulligan D., Bologna R. Botulinum toxin: historical perspective and treatment of neurogenic and idiopathic overactive bladder // Therapy. 2009. Vol. 6, № 2. P. 165175.
- Smith C.P., Vemulakonda V.M., Kiss S., Boone T.B., Somogyi G.T. Enhanced ATP release from rat bladder urothelium during chronic bladder inflammation: effect of botulinum toxin A // Neurochem Int. 2005. Vol. 47, № 4. Р. 291-297.
- Myung S.C., Song E.S., Keum E.M., Lee M.Y., Kim K.D. The Effects of Botulinum Toxin-A on Calcium Channel and KDR Channel of the Detrusor Muscle in Rat Bladder Body // Korean J. Urol. 2004. Vol. 45, №6. Р. 585-591.
- Mossa H.E., Burnstock G. A comparative study of electrical field stimulation of the guinea-pig, ferret and marmoset urinary bladder // European Journal of Pharmacology. 1985. Vol. 114, № 27. P. 311-316.
