







ВВЕДЕНИЕ
Традиционно считают, что «мишенью» гексанового экстракта Serenoa repens (HSRE) является собственно предстательная железа, где препарат демонстрирует возможность ингибиции 5α-редуктазы, а также антиандрогенный, антипролиферативный и противовоспалительный эффекты [1–4]. Однако в литературе встречаются данные о том, что терапевтическое действие HSRE может осуществляться и на уровне мочевого пузыря.
Нами была высказана гипотеза о прямом действии HSRE на обструктивный мочевой пузырь, препятствующем развитию его дисфункции [5]. В ее подтверждение было проведено исследование на белых беспородных крысах-самках, в котором показано, что в условиях тяжелой инфравезикальной обструкции (ИВО) применение HSRE, по сравнению с группой без терапии, приводило к следующим эффектам:
- развитию более выраженной гипертрофии мочевого пузыря и «задержке» на этой стадии развития инфравезикальной обструкции (ИВО), тогда как у нелеченых животных наблюдали декомпенсацию детрузора;
- сохранению, в большей степени, таких функциональных параметров мочевого пузыря, как комплаентность и эвакуаторная способность;
- предупреждению формирования гиперактивности детрузора, которую наблюдали в контрольных опытах;
- лучшей сохранности гладкомышечных клеток детрузора, что косвенно проявилось в более низкой активности креатинфосфокиназы в крови;
- уменьшению степени воспаления и склерозирования стенки мочевого пузыря при гистологическом исследовании;
- менее выраженным нарушениям показателей функционального состояния почек: скорости клубочковой фильтрации и активности канальцевой реабсорбция натрия.
В целом, терапия HSRE способствовала более высокой выживаемости животных. Поскольку опыты были проведены на самках крыс, то выявленные положительные влияния HSRE были связаны с непосредственным действием препарата именно на мочевой пузырь и не опосредованы потенциальным влиянием на предстательную железу [6].
Механизмы влияния HSRE на мочевой пузырь в настоящий момент не ясны. В данной статье мы представляем гипотезу о возможных механизмах действия гексанового экстракта Serenoa repens на функцию мочевого пузыря при ИВО на основе анализа литературных и собственных данных.
МАТЕРИАЛЫ И МЕТОДЫ
Проанализированы данные литературы из баз PubMed и eLibrary.ru за 25-летний период, связанные с изучением патогенеза дисфункции мочевого пузыря при развитии ИВО, клеточных и молекулярных механизмов, посредством которых реализуются патологические и адаптационно-приспособительные процессы в клетках мочевого пузыря, а также влияние на эти процессы HSRE.
РЕЗУЛЬТАТЫ
Влияние HSRE на динамику развития дисфункции мочевого пузыря при ИВО
Установлено, что частичная ИВО вызывает морфологические изменения в мочевом пузыре, а также изменение его функции [7]. Повышение внутрипузырного давления во время опорожнения мочевого пузыря при ИВО – патогномоничный уродинамический фактор, являющийся «первичным» по отношению к ряду вызываемых им патофизиологических последствий в виде модуляции сигнальных путей, участвующих в регуляции клеточного метаболизм, а также к последующим гистологическим изменениям, происходящим почти во всех клеточных компартментах: уротелии, субуротелии, гладкомышечных клетках детрузора, экстрацеллюлярном матриксе и нейронных структурах [8]. Критическим фактором при ИВО выступает тканевая гипоксия, развивающаяся из-за дисбаланса между повышенной потребностью в кислороде и его более низким поступлением вследствие ухудшения микроциркуляции, вызванного стойким повышением внутрипузырного давления и опережающим увеличением массы детрузора по отношению к неоангиогенезу, что запускает каскад патологических реакций уже на ранних стадиях ремоделирования мочевого пузыря, вызванного ИВО [9].
Развивающаяся при ИВО дисфункция мочевого пузыря, по мнению ряда авторов, претерпевает три последовательные стадии: гипертрофии детрузора, компенсации и декомпенсации, тогда как другие авторы выделяют лишь две фазы патологического процесса – компенсации и декомпенсации [10, 11]. В компенсированной фазе мышца детрузора увеличивает силу сокращений, чтобы преодолеть возросшее сопротивление, что ведет к гипертрофии миоцитов и сохранению эвакуаторной функции органа [11]. При этом происходит стимуляция ангиогенеза для обеспечения кровоснабжения возрастающей массы мочевого пузыря, что на определенном этапе обеспечивает адаптацию органа к дефициту кислорода [10, 12]. Развивающаяся гипертрофия детрузора сопровождается увеличением массы органа и характерными гистологическими изменениями мышечной оболочки мочевого пузыря [13].
S.A. Zderic и соавт. декларировали, что начальная стадия развития дисфункции мочевого пузыря при ИВО характеризуется следующими признаками: (1) прогрессирующим увеличением массы мочевого пузыря, (2) потерей реакции на электростимуляцию, (3) потерей реакции на стимуляцию прямым агонистом и (4) уменьшением способности полностью эвакуировать мочу. Однако в большинстве случаев за счет включения адаптивных процессов следует период относительной компенсации, в течение которого детрузор может частично или полностью восстанавливать сократительную активность в ответ на электростимуляцию или стимуляцию агонистом [14]. При длительном существовании и/или прогрессировании ИВО может наступать окончательная функциональная декомпенсация гипертрофированного мочевого пузыря в результате дистрофических изменений миоцитов и интрамуральных нейронов с частичной их утратой и замещением соединительной тканью [15, 16]. При этом эвакуаторная функция мочевого пузыря прогрессивно ухудшается, рост массы мочевого пузыря и ангиогенез прекращаются. Конец этой фазы характеризуется преобладанием фиброзной ткани в стенке мочевого пузыря за счет замещения внеклеточным матриксом мышечных компонентов, снижением емкости мочевого пузыря и его плохой податливостью (комплаентностью) [7, 16–18]. Этот принцип проиллюстрирован на рисунке 1.
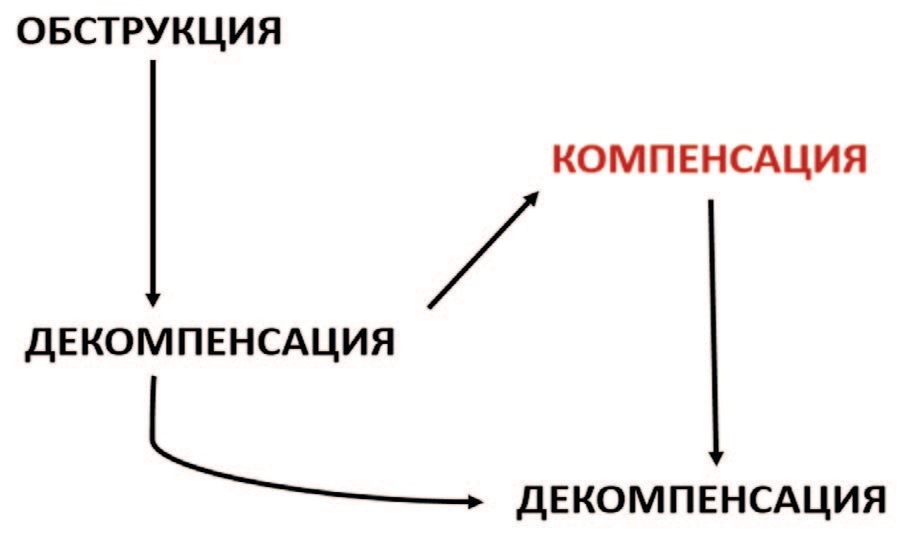
Рис. 1. Схематическое представление фаз компенсации и декомпенсации при экспериментальной ИВО [14]
Fig.1. Schematic representation of compensation and decompensation phases in experimental IVO [14]
Наряду со склерозированием мочевого пузыря происходит деградация интрамуральных нервных структур, что ведет к утрате нейрогенной регуляции сокращений с формированием гиперактивности детрузора, характеризующейся спонтанными сокращениями, не связанными с мочеиспусканием, с повышенной частотой мочеиспускания или без ее увеличения и последующим уменьшением объема мочеиспускания и емкости мочевого пузыря [19–21]. Именно эти изменения мы наблюдали в контрольной серии опытов нашего исследования [6].
Способность компенсации функции мочевого пузыря при ИВО важна для понимания потенциальной возможности медикаментозного протективного воздействия на этапе обратимых изменений. Именно эту возможность продемонстрировали проведенные нами исследования по влиянию HSRE на гипертрофированный мочевой пузырь [6]. В условиях выраженной ИВО мы наблюдали выраженную гипертрофию детрузора у животных основной группы, что характерно для описанной выше первой стадии патологического процесса компенсации – гипертрофии. В то же время состояние мочевого пузыря у крыс контрольной группы можно характеризовать как декомпенсированное, поскольку, несмотря на гипертрофию детрузора, все показатели функционального состояния мочевого пузыря значительно ухудшились, а в стенке мочевого пузыря наблюдали выраженное воспаление и склероз. То есть назначение HSRE, вероятно, затормозило развитие патологического процесса на этапе гипертрофии – начале фазы декомпенсации, по сравнению с контрольной группой, где развилась выраженная декомпенсация мочевого пузыря (рис. 2).
![Схема патогенеза ИВО и возможного влияния HSRE [адаптировано по 18]](https://ecuro.ru/sites/default/files/issue/2023_2/16,2.jpg)
Рис. 2. Схема патогенеза ИВО и возможного влияния HSRE [адаптировано по 18]
Fig.2. Schematic representation of IVO pathogenesis and potential effects of HSRE [adapted from 18]
Возможное влияние HSRE на адаптивные и дезадаптивные звенья патогенеза дисфункции гипертрофированного мочевого пузыря при ИВО
Адаптивные реакции могут противодействовать развитию патологического процесса, в том числе при ИВО, только в течение ограниченного периода времени, и при неустраненном патологическом факторе эти реакции могут не только утратить эффективность, но и перейти в категорию патологических. Определенные медикаментозные воздействия могут способствовать сохранению адаптивного характера метаболических реакций более длительное время, воздействуя на определенные звенья патогенеза. Учитывая установленное многофакторное действие HSRE на метаболизм клеток гиперплазированной предстательной железы, мы проанализировали возможное действие этого препарата на разные звенья патогенеза дисфункции мочевого пузыря при ИВО.
Противовоспалительное действие
Антиоксидантное действие
Основным последствием хронической гипоксии, развивающейся при ИВО, является оксидантный стресс вследствие избыточной продукции митохондриями активных форм кислорода (ROS – reactive oxygen species), таких как супероксид-анион радикалы и гидроксильные радикалы, вызывающие повреждение клеточных структур и развитие асептического воспаления. Опубликованы данные об увеличении выработки ROS и малонового диальдегида (MDA – конечного продукта свободно-радикальных реакций) в мочевом пузыре у крыс с тяжелой ИВО [16, 22–25]. В то же время активность таких антиоксидантных ферментов, как супероксиддисмутаза и каталаза, в мочевом пузыре уменьшается, что свидетельствует о снижении антиоксидантной способности ткани при ИВО [26]. Оксидантное повреждение клеток мочевого пузыря приводит к его дисфункции со снижением сократительной способности миоцитов детрузора и эвакуаторной функции мочевого пузыря, а также развитием гиперчувствительности и нестабильности детрузора за счет повреждения интрамуральных нервных структур и нарушения нейрогенной регуляции сокращений [24, 27–30]. Следовательно, ROS могут быть патогенным фактором воспаления дисфункционального детрузора, а антиоксиданты, вероятно, быть полезны для лечения дисфункции мочевого пузыря, вторичной по отношению к обструкции [31].
Именно таким эффектом обладает HSRE. Было отмечено, что экстракт вееролистной пальмы Serenoa repens является богатым источником фенольных соединений (фенолов и флавоноидов), обладающих потенциальной антиоксидантной, противомикробной и противовоспалительной активностью. Заявлено, что эти фенольные соединения показали высокую активность в отношении свободных радикалов [32]. Исследовано влияние Serenoa repens на маркеры окислительного стресса, воспаления и факторы роста у крыс с ожирением и гиперплазией предстательной железы, индуцированной тестостероном. Применение HSRE привело к значительному уменьшению массы предстательной железы, уровней нитритов и МDА (маркеры оксидантного стресса); повышению содержания глутатиона, активности антиоксидантных ферментов (супероксиддисмутазы и каталазы), а также значительному уменьшению воспалительных факторов TNF-α, IL-1β, IL-6 и факторов роста (VEGF и FGFb). То есть HSRE продемонстрировал высокую антиоксидантную и противовоспалительную активность, способность модулировать воспалительные реакции в предстательной железе. В этой экспериментальной работе подтверждена антиоксидантная активность Serenoa repens: снижение перекисного окисления липидов и предотвращение дальнейшего повреждения тканей ROS. Однако этот эффект авторы наблюдали в предстательной железе и связывали, в первую очередь, с антиандрогенным эффектом HSRE [33]. Тем не менее, мы предполагаем, что протективное действие HSRE на мочевой пузырь при ИВО потенциально может быть реализовано, в том числе, и через механизм антиоксидантной активности.
Воздействие на систему провоспалительных цитокинов
Оксидантному повреждению клеточных стриктур мочевого пузыря при ИВО неизбежно сопутствует развитие асептического воспаления через активацию синтеза ряда сигнальных молекул, в том числе за счет молекулярных агентов, высвобождаемых из умирающих клеток. Эти, так называемые молекулярные паттерны, «связанные с опасностью» (Danger Associated Molecular Patterns – DAMPs), распознаются Nod-подобными рецепторами (NLRs), образующими многомерные структуры, называемые инфламмасомами, которые способствуют образованию и высвобождению провоспалительных цитокинов-интерлейкинов IL-1β и IL-18. Было показано присутствие NLRs в уротелии и их способность реагировать на такие вещества, как АТФ и урат натрия, которые являются обычными компонентами мочи, и повышение экспрессии IL-1β уже на ранней стадии формирования ИВО [34–36]. Развивающееся при ИВО воспаление приводит к формированию симптомов «раздражения» (фазы накопления), таким как частое мочеиспускание, ургентность и недержание мочи, которые часто более значимы для качества жизни пациентов, чем первоначальные обструктивные симптомы. Противовоспалительная терапия может предотвратить или замедлить прогрессирование дисфункции мочевого пузыря при частичной ИВО [37–40].
Vela Navarette и соавт. описали влияние HSRE на воспалительный статус предстательной железы в клиническом исследовании, показавшем значительное и достоверное снижение количества В-лимфоцитов и провоспалительных цитокинов (TNFα и IL-1β) на фоне терапии [41]. Примечательно, что HSRE, по-видимому, влияет на множественные функциональные пути воспаления не только через семейство цитокинов, но и через сигнальные пути глюкокортикоиды/PPAR (peroxisome proliferator-activated receptor), MAPK (mitogen-activated protein kinase), суперсемейство TNF и пути COX/LOX. Некоторые из этих генов кодируют молекулы, которые, как предполагают, являются ключевыми медиаторами в процессе воспаления, такими как IL6 и IL8 в стромальных клетках или IL1α, IL1β и их рецептор IL1R1 в эпителиальных клетках [42-45]. В частности, положительное влияние HSRE на экспрессию ряда генов, кодирующих синтез провоспалительных цитокинов, в частности гена IL1B описано в работах N. Sirab и соавт. и A. Latil и соавт. [46, 47]. Ранее нами также было показано снижение уровня IL-1β через 3 и 6 месяцев лечения экстрактом Serenoa repens больных хроническим простатитом [48].
Еще один путь формирования воспалительной реакции в мочевом пузыре при ИВО посредством цитокинов реализуется с участием фактора, ингибирующего миграцию макрофагов (MIF) – Т-клеточного цитокина, который является ключевым медиатором врожденного иммунитета и плейотропным воспалительным цитокином, играющим важную роль в воспалительном процессе и рекрутинге лейкоцитов [49–51]. MIF экспрессируется как периферически, так и центрально в различных клетках, таких как макрофаги, эпителий мочевыводящих путей, толстой кишки, в нейронах спинного мозга, головного мозга и глии [52, 53] и высвобождается из предварительно сформированных пулов клеток при иммунных реакциях и/или травме нервной системы [54].
Уротелий представляет собой богатый источник MIF, который является важным медиатором воспаления мочевого пузыря [55]. Также показано, что MIF конститутивно экспрессируется на высоких уровнях в тучных клетках, в том числе человека, количество которых в слизистой и подслизистой оболочках мочевого пузыря крыс увеличивается при воспалении и частичной ИВО [56, 57]. Считают, что в условиях воспаления MIF способствует пролиферации и выживанию фибробластов, синтезу коллагена и деградации мышечной ткани, то есть усиливает процесс склерозирования мочевого пузыря [58, 59], а блокада его активности (у мышей с нокдауном гена MIF), значительно ингибирует профиброзную активность [60]. Это подтверждено в опытах на генно-модифицированных мышах и в опытах in vitro на культурах клеток [61]. Другие авторы также подтверждают, что MIF участвует в последовательности событий, приводящих к потере мышечных клеток детрузора и их замещению соединительной тканью при ИВО, а стратегии, направленные на противодействие синтезу, высвобождению или биологической активности MIF, могут предотвратить или отсрочить гипоактивность детрузора и задержку мочи [49].
Терапевтическое действие HSRE может реализоваться и через влияние на этот регуляторный путь. В исследовании A. Latil и соавт. показано, что экспрессия MIF на фоне применения HSRE у больных ДГПЖ существенно снижается, что сочетается с положительным клиническим эффектом терапии [47]. Снижение экспрессии MIF может являться следствием общего противовоспалительного действия HSRE и реализоваться опосредованно через гипорегуляцию HIF1А [62].
С учетом универсальности механизмов развития воспаления и неспецифичности противовоспалительного действия HSRE, описанные эффекты регуляции провоспалительных цитокинов, включая IL-1β и MIF, могут играть роль и при купировании воспаления мочевого пузыря при ИВО.
Воздействие на синтез простагландинов
Еще одним потенциальным механизмом развития воспаления мочевого пузыря при ИВО является активация «каскада арахидоновой кислоты» с усилением синтеза простагландинов (PG), в первую очередь простагландина Е2 (PGE2). Наряде с тем, что PGE2 является эндогенным модулятором функции мочевого пузыря в нормальных физиологических условиях, при его избыточном синтезе он может стать участником патофизиологических процессов. В норме PG, в том числе PGE2, содействуют поддержанию тонуса гладкой мускулатуры нижних мочевыводящих путей и обладают вазодилатирующим действием, способствуя регуляции притока крови к мочевому пузырю [63–67], за счет чего, по данным ряда авторов, обладают цитопротективным действием при ИВО [68].
Образование PGE2 происходит локально в мышцах и слизистых оболочках мочевого пузыря в ответ на многочисленные раздражители, такие как растяжение, стимуляция нервов, травма и воспаление [65–67]. Показано, что чрезмерное растяжение мочевого пузыря из-за обструкции уретры активирует синтез PG у собак, крыс и человека [66, 69, 70] и это сочетается с изменением функционального состояния и морфологии мочевого пузыря [71].
Синтез PGE2 осуществляется из арахидоновой кислоты при участии фермента циклоксигеназы-2 (COX-2), экспрессия которого повышается при формировании обструкции мочевого пузыря у крыс, а также в ответ на растяжение клеток мочевого пузыря in vitro. Установлено, что уровень мРНК COX-2 повышается на фоне частичной обструкции уретры [72, 73].
Увеличению синтеза PG способствует также повышение концентрации в клетках свободной арахидоновой кислоты, являющейся предшественником PG, при развитии обструкции мочевого пузыря [74]. Это обусловленно ее повышенным освобождением из фосфолипидов клеточных мембран под действием фосфолипазы А2, активируемой в условиях оксидативного стресса и окисления мембранных фосфолипидов активными формами кислорода [75]. Есть данные, что повышенный синтез PG вследствие активации COX-2 при ИВО может быть вовлечен в развитие гипертрофии мочевого пузыря, а также гиперактивности детрузора [65, 73]. Внутрипузырное введение PGE2 крысам вызывает гиперактивность детрузора и стимулирует рефлекторное мочеиспускание [76], а у людей – к гиперактивности детрузора, снижению емкости мочевого пузыря и ургентности [77].
A. Schroder и соавт. считают, что эндогенный PGE2 и его рецептор EP1 не играют значимой роли в модуляции мочеиспускания как при нормальных обстоятельствах, так и при гипертрофии детрузора в результате ИВО, но опосредуют возникновение детрузорной гиперактивности в патологических условиях при ИВО [78]. Стимуляция рецептора EP1 играет ключевую роль в развитии гиперактивности детрузора при ИВО у мышей за счет усиления афферентных сигналов с рецепторов мочевого пузыря в ЦНС [78–80]. Ингибирование передачи сигналов по пути «каскада арахидоновой кислоты» может быть терапевтически полезным при лечении симптомов мочевого пузыря, связанных с обструкцией. Ранее была показана дозозависимая способность HSRE вмешиваться в метаболизм каскада арахидоновой кислоты и ингибировать ключевые ферменты (фосфолипазу А2, липоксигеназу и циклоксигеназу), тем самым определяя торможение синтеза простагландинов и лейкотриенов [81–83]. В исследовании A. Latil и соавт. также было показано позитивное влияние HSRE на экспрессию генов, связанных с метаболизмом арахидоновой кислоты: PTGES2, PTGES3 – кодируемые белки которых представляют собой варианты мембраносвязанной простагландин Есинтазы, преобразующей эндопероксид простагландина H2 (PGH2) в простагландин E2 (PGE2), а также PTDS2, кодирующий циклоксигеназу [47]. На модуляцию HSRE гена PTGS2 ранее указали N. Sirab и соавт. [46].
Таким образом, вероятна реализация противовоспалительного действия HSRE при частичной ИВО через влияние на каскад арахидоновой кислоты и снижение синтеза простагландинов. Кроме того, с учетом установленной роли последних в возникновении гиперактивности детрузора, можно предположить, что воздействие HSRE на симптомы фазы накопления и гиперактивность мочевого пузыря реализуется и через этот механизм.
Регулирующее влияние HSRE на клеточные адаптационные процессы
Помимо ингибирующего влияния на звенья патогенеза дисфункции мочевого пузыря при ИВО, HSRE может оказывать терапевтическое действие путем влияния на адаптивные клеточные механизмы, направленные на сохранении функциональной активности клеток и их выживаемость.
Влияние на реакции, опосредованные фактором, индуцируемым гипоксией
Один из таких механизмов реализуется с участием фактора, индуцируемого гипоксией (HIF) [84]. Комплекс факторов транскрипции HIF состоит из α – и βсубъединиц [85, 86]. Ряд клинических и экспериментальных исследований демонстрируют, что ИВО запускает индуцированную гипоксией гиперрегуляцию фактора HIF-1α – основного регулятора транскрипции клеточного ответа на гипоксию при снижении концентрации кислорода в ткани до 1% [87]. И наоборот, HIF-1β – ядерный транслокатор рецептора арильных углеводородов (aryl hydrocarbon receptor nuclear translocator – Arnt) – управляется независимым от кислорода способом [88]. Еще одна субъединица HIF, а именно HIF-2β (Arnt2), может образовывать гетеродимеры, способные к трансактивации, с HIF-1α [89].
H.A. Stephany и соавт. сообщили об активации HIF-1 и повышении уровней мРНК Arnt2 у мышей с растяжением мочевого пузыря. При гипоксии субъединицы HIF1α стабилизируются путем образования комплексов с HIF-1β/Arnt, что представляет собой прототипный путь трансактивации, опосредованной HIF-1. Arnt2 может связываться с субстанциями, чувствительными к гипоксии, в качестве адаптивной реакции на низкое напряжение кислорода в мочевом пузыре [90].
G. Vangeison и соавт. заявили о роли HIF-1α в сохранении жизнеспособности нейронов при ИВО: более высокие уровни HIF-1α в группе с умеренной обструкцией могут предотвращать нарушения нейрогенно-обусловленных сократительных реакций [91]. Однако в других работах, где была использована модель сильно выраженной ИВО, не отметили никаких изменений в уровнях HIF-1α у крыс до 4 недель наблюдения [37], что позволило предположить, что гипоксия может играть потенцирующую роль именно на ранних стадиях ДГПЖ, после чего этот путь адаптации становится неэффективным [27].
M. Ekman и соавт. при тяжелой ИВО продемонстрировали гиперрегуляцию Arnt2, но не HIF-1α, как на уровне мРНК, так и белка, а экспрессия фактора транскрипции HIF-2β/Arnt2 оказалась повышена. При этом у крыс с умеренной ИВО зарегистрирована повышенная экспрессия генов и белка HIF-1α и HIF2β/Arnt2 [86].
Таким образом, представляется, что воздействие на сигнальный путь, реализуемый с участием HIF-1α и Arnt2, может иметь перспективы в плане профилактики расстройств функции мочевого пузыря при ИВО, в частности нейрогенной регуляции сокращений, приводящих к формированию гиперактивности детрузора. В нашем исследовании [6] было продемонстрировано, что терапия HSRE приводит к снижению гиперактивности детрузора при искусственно вызванной ИВО у крыс [6].
Влияние на транскрипцию генов, регулирующих адаптивные реакции
Адаптация к гипоксии и повреждению тканей мочевого пузыря при обструкции мочевого пузыря также происходит путем активации транскрипции различных генов [92]. Ядерный фактор-2, связанный с эритроидным ядерным фактором (Nrf2 – Nuclear factor, erythroid 2-like 2), и ядерный фактор «каппа би» (NF-κB) являются двумя наиболее важными факторами транскрипции. Между этими двумя критическими путями существует функциональное перекрестное взаимодействие. Nrf2 активируется в условиях окислительного стресса и модулирует клеточные реакции на окислительный стресс [93].
Продемонстрировано, что при ИВО в гладкомышечных клетках обструктивного мочевого пузыря повышается активность NF-κB, контролирующего экспрессию генов иммунного ответа, апоптоза и клеточного цикла [94]. Установлено, что окислительный стресс может индуцировать транслокацию NF-κB в ядро, что наблюдали в опытах с тяжелой ИВО на фоне двукратного повышения уровня малонового диальдегида (MDA) – маркера перекисного окисления липидов [27, 94–96]. В состоянии покоя NF-κB изолирована в цитоплазме клетки посредством ингибитора IkBα, а его транслокация в ядро обусловлена окислительной деградацией этого ингибитора, где он включает экспрессию специфических генов [97, 98]. В частности, установлено, что NF-кB1 является ДНКсвязывающей субъединицей белкового комплекса транскрипционного фактора NF-κB [47].
Исследования показали, что экспрессия мРНК и белка Nrf2, оказывающего цитопротекторное действие в мочевом пузыре, была значительно повышена у крыс с тяжелой обструкцией [24]. С учетом перекрестной связи этих факторов транскрипции, ядерная транслокация NF-κB может быть ограничена усилением регуляции цитопротекторного Nrf2 в мочевом пузыре [27]. Таким образом, различные степени обструкции и гипоксии изменяют функцию мочевого пузыря и молекулярные реакции с участием факторов NF-κB и Nrf2 [97, 98]. Авторы предположили, что ингибирование активности NF-κB может уменьшить воздействие окислительного стресса при ИВО [27]. Применение ингибитора NF-κB на модели крыс с ишемически-реперфузионным повреждением защитило мочевой пузырь от окислительного стресса [98].
N. Sirab и соавт. установили, что HSRE модулирует экспрессию связанных с воспалением генов, включая NF-кB1 и NF-kBIZ, с последующим торможением синтеза провоспалительных цитокинов [46]. A. Latil и соавт. также указали на позитивное влияние HSRE на экспрессию ряда генов, связанных с оксидативным стрессом и воспалением, в том числе HIF1A и NF-кB1 [47]. Это указывает на возможное наличие дополнительных защитных механизмов действия HSRE, связанных с влиянием на сигнальные пути, регулируемые HIF-1 и факторами транскрипции NF-κB / Nrf2 при частичной ИВО.
Влияние HSRE на факторы роста и пролиферации клеток
В нашем эксперименте было показано, что терапия HSRE способствует более выраженной гипертрофии мочевого пузыря, адаптирующей его к повышенной нагрузке. Этот процесс регулируется факторами роста и пролиферации [99]. Установлено, что при ИВО активируется синтез ряда факторов роста, в частности фактора роста фибробластов (FGF), трансформирующего фактора роста β (TGF-β), фактора роста эпидермиса (EGF) и фактора роста нервов (NGF), что опосредовано развитием воспалительной реакции с миграцией в ткани лимфоцитов и активацией синтеза ряда цитокинов (IL-1, IL-6, TNF-α) [99].
Семейство факторов роста фибробластов (FGF) участвует при нормальном росте мочевого пузыря, а его кислая (aFGF) и щелочная/основная (bFGF) формы синтезируются в ткани мочевого пузыря. Рецепторы FGF находятся на фибробластах слизистой оболочки мочевого пузыря и серозной оболочки. Известна роль bFGF в инициации и стимулировании пролиферации фибробластов, в том числе при развитии гипертрофии мочевого пузыря [100]. Экспрессия bFGF увеличивается в обструктивном мочевом пузыре и может быть фактором, влияющим на рост массы мочевого пузыря при ИВО [101]. Однако bFGF также способен стимулировать пролиферацию фибробластов с повышением потенциала синтеза коллагена, что может вести к снижению податливости (комплаентности) мочевого пузыря и к появлению симптомов нарушенного мочеиспускания [101].
Другим регулятором клеточной пролиферации является трансформирующий фактор роста-β (TGF-β) – пептид, который чаще всего идентифицируется как агент, подавляющий клеточный рост или фактор клеточной дифференцировки, что связано с его влиянием на синтез белков клеточного матрикса и способностью модулировать секрецию других факторов роста. TGF-β синтезируется мочевым пузырем взрослого человека, и его экспрессия в мочевом пузыре изменяется при ИВО. Показано, что соотношение этих факторов роста при развитии ИВО существенно меняется. В первые часы (до 48 часов) после возникновения частичной обструкции экспрессия мРНК bFGF в мочевом пузыре значительно повышается, тогда как экспрессия мРНК TGF-β в ранний период формирования ИВО подавлялась, снижаясь до 50% от контрольных уровней [101]. Временное повышение экспрессии bFGF при снижении экспрессии TGF-β согласуется с ранней фазой гипертрофического роста при ИВО, включающей стимулированную пролиферацию как фибробластов слизистой и серозной оболочек мочевого пузыря, так и роста гладкомышечных клеток [99].
Важно, что изменения на ранних стадиях ИВО носят еще обратимый характер: при устранении обструкции через 2 недели экспрессия bFGF в мочевом пузыре быстро снижалась, а экспрессия TGF-β нарастала. Таким образом, когда стимул к гипертрофии мочевого пузыря был ослаблен, паттерн экспрессии факторов роста был обращен вспять и, в ответ, масса мочевого пузыря снижалась, приближаясь к норме в результате механизма клеточной гибели – апоптоза [99, 100].
Подобные изменения в профиле этих факторов клеточной пролиферации выявлены в других исследованиях, где было показано, что при чрезмерном временном перерастяжении мочевого пузыря кроликов или искусственно вызванной частичной ишемии мочевого пузыря, также происходило повышение экспрессии мРНК bFGF и снижение экспрессии мРНК TGF-β, что указывало на связь этих изменений с развитием ишемии/гипоксии мочевого пузыря при ИВО [102, 103]. Учитывая, что bFGF является мощным ангиогенным и пролиферативным фактором, можно предположить, что повышение экспрессии bFGF, стимулированное гипоксией в ткани гипертрофированого мочевого пузыря при ИВО, является результатом паракринного стимула к восстановлению нарушенной микроциркуляции в органе (рис. 3).
![Схема влияния растяжения стенки мочевого пузыря на доступность EGF к эпителиальным клеткам [Адаптировано по 107]](https://ecuro.ru/sites/default/files/issue/2023_2/16,3.jpg)
Рис. 3. Схема влияния растяжения стенки мочевого пузыря на доступность EGF к эпителиальным клеткам [Адаптировано по 107]
Fig 3. Influence of bladder wall distension on EGF availability to epithelial cells [Adapted from 107]
Фактор роста эпидермиса (EGF) – пептид, способный влиять на деление эпителиальных клеток в уротелии мочевого пузыря. Хотя нет доказательств того, что EGF образуется в ткани мочевого пузыря, он в изобилии секретируется в мочу клетками почек, преимущественно дистальным канальцевым эпителием [104, 105]. Факт быстрой пролиферативной реакции уротелия в ответ на обструкцию и его чрезвычайной чувствительности к стимуляции EGF позволил предположить, что этот фактор роста играет роль при развитии гипертрофической реакции [99]. Рецептор EGF (EGFr) представляет собой трансмембранный белок, в отношении которого наблюдается разительная градация клеток уротелия. Иммуногистохимические исследования показали, что базальноклеточный слой уротелия сильно обогащен этими рецепторами, тогда как в более дифференцированных промежуточных клетках EGFr их мало, а высокодифференцированные поверхностные (зонтичные) клетки вообще не имеют их [106]. Поскольку зонтичные клетки образуют относительно непроницаемый барьер для мочи, при нормальных условиях EGF мочи не получает доступ к своим многочисленным рецепторам на базальных уротелиальных клетках. Нарушение барьерной функции уротелия мочевого пузыря в результате его чрезмерного растяжения ведет к проникновению EGF с мочой к базальным клеткам уротелия и последующему чрезмерному росту этого слоя (рис. 3) [99].
Выраженную гиперплазию уротелиальной выстилки гипертрофированного мочевого пузыря мы наблюдали и в наших экспериментах [6]. Таким образом, серия экспериментов с участием различных моделей частичной ИВО на животных демонстрирует, что происходит обширное ремоделирование ткани мочевого пузыря. Представленные данные указывают на то, что изменения экспрессии bFGE, EGF и TGF-β являются важными компонентами этого раннего ответа, действующего аутокринным или паракринным образом для стимуляции роста мочевого пузыря. EGF также может играть роль в этом процессе благодаря высокой концентрации в моче и нарушению обычно непроницаемого уротелиального барьера при ИВО.
Важно, что первая фаза изменений мочевого пузыря при ИВО обратима после устранения обструкции: экспрессия факторов роста возвращается к контрольному уровню, когда мочевой пузырь возвращается к своим нормальным параметрам.
Терапия HSRE может влиять на регуляцию пролиферации клеток при ИВО. М. Paubert-Braquet и соавт. изучали влияние HSRE на стимулируемую bFGF и EGF пролиферацию клеток предстательной железы, полученных из биопсийного материала ткани гиперплазированной предстательной железы человека. EGF и bFGF индуцировали выраженную пролиферацию клеток предстательной железы человека в культуре in vitro, тогда как HSRE ингибировал эту пролиферацию дозозависимым образом. При этом HSRE и его компоненты оказывали большее влияние на bFGF-сигнальную индукцию клеточной пролиферации [3, 108]. В экспериментальном исследовании на крысах J. ColadoVelázquez III и соавт. продемонстрировали достоверное снижение активности факторов роста bFGF и VEGF на фоне приема HSRE [33].
То есть, с учетом опубликованных данных, имеется вероятность того, что влияние SRE на обструктивный мочевой пузырь может реализоваться и через регуляцию факторов роста, в первую очередь bFGF, TGF-β и EGF.
Регулирующее действие HSRE на нейрогенные структуры мочевого пузыря
Сократительная способность гладкомышечных структур мочевых путей является одной из основных целей фармакотерапии СНМП, поскольку их чрезмерное сокращение в предстательной железе способствует развитию функциональной ИВО, а неконтролируемые сокращения в мочевом пузыре – гиперактивности детрузора. При этом в значительной степени сократительная активность мочевого пузыря регулируется нейрогенными механизмами, которые нарушаются при ИВО, в том числе за счет дегенерации интрамуральных нервных структур. В поддержании их жизнеспособности и функции участвует фактор роста нервов (NGF). Поскольку этот пептид обычно секретируется в небольшом количестве, его непрерывный синтез позволяет поддерживать соответствующую концентрацию в большинстве тканей взрослого человека. Экспрессия мРНК NGF возрастает в мочевом пузыре человека и крыс при ИВО, что свидетельствует о компенсаторном характере этой реакции в ответ на повреждение нейронов, и частично восстанавливается после устранения обструкции уретры [109]. Значительная гипертрофия афферентных и эфферентных нейронов в тазовых ганглиях крыс с ИВО является еще одним следствием повышенной экспрессии и высвобождения NGF в паренхиматозных клетках гипертрофированного мочевого пузыря. С этой точки зрения, гиперактивность мочевого пузыря, наблюдаемая при ИВО, может быть результатом избыточной выработки NGF в мочевом пузыре в острой фазе после возникновения обструкции. Дисфункции мочевого пузыря при ИВО также способствует изменение в рецепторном аппарате гладкомышечных клеток с нарушением баланса адренергических и холинергических звеньев регуляции [5].
Считают, что активные жирные кислоты, присутствующие в HSRE, способны оказывать существенное расслабляющее действие на сокращения гладкомышечных элементов тканей животных, в том числе ПЖ [110, 111]. Рядом исследований показано, что SRE обладает свойствами ингибирования α1-адренорецепторов [112], М-холинорецепторов [113, 114] и спазмолитической активностью [115]. Некоторые компоненты SRE могут ингибировать вызванные адреномиметиком фенилэфрином сокращения семявыносящего протока, то есть обладают α-адреноблокирующим действием [116].
В отдельных экспериментальных работах SRE проявил значительную связывающую активность в отношении рецепторов автономной нервной системы: α1-адренергических и мускариновых рецепторов нижних мочевых путей крыс, включая как предстательную железу, так и мочевой пузырь. SRE препятствовал in vitro связыванию Н3-празозина (антагонист α-адренорецепторов) и Н3-N-метилскополамина (антагонист М-холинорецепторов), не влияя на связывание αβМе-АТФ (антагонист пуринергических рецепторов) с тканями предстательной железы и мочевого пузыря, в дозозависимом отношении, что свидетельствует о его способности блокировать α1-адренорецепторы и М-холинорецепторы не только в ПЖ, но и в мочевом пузыре. При этом способность к связыванию с М-холинорецепторами, которые широко представлены в детрузоре, была в 2–4 раза выше, чем с α-адренорецепторами. Установлено, что SRE блокирует 45% мест связывания для Н3-празозина и Н3-N-метилскополамина как в ПЖ, так и в мочевом пузыре [113, 114].
В опытах in vivo при пероральном введении SRE крысам в течение 4 недель выявили значительное снижение мест связывания М-холинорецепторов в мочевом пузыре и в слюнной железе (как орган сравнения), причем эффект SRE на холинорецепторы мочевого пузыря проявлялся в значительно (10 раз и более) меньших дозах препарата, чем на рецепторы слюнной железы, что свидетельствует о большей тропности экстракта к рецепторам клеток мочевого пузыря. В то же время количество мест связывания α-адренорецепторов в ПЖ при дозе препарата 6 мг/кг в сутки возрастало [113‑117].
Имеются указания на то, что мышечная релаксация, вызываемая липидным SRE, связана с экструзией Ca++ посредством Na+/Ca++ обменника, как было описано для других липидных соединений. Обменник Na+/Ca++ – это трансмембранный белок цитоплазматической мембраны, транспортирующий ионы кальция из клетки в обмен на ионы натрия, поступающие в клетку, активность которого регулируется химическими взаимодействиями с липидами из липидного бислоя. Липидный SRE, который сильно обогащен жирными кислотами, может влиять на Na+/Ca++ обменник вторично по отношению к изменениям свойств клеточной мембраны [118], в том числе путем, опосредованным цАМФ (циклическим аденозинмонофосфатом). Путь цАМФ важен для расслабления гладких мышц при опосредованных а1-рецептором сокращениях гиперплазированной ткани предстательной железы [119].
Таким образом, SRE может способствовать расслаблению гладкомышечных структур с помощью плейотропных механизмов, в конечном счете влияющих на сократительный аппарат. Активация Na+/Ca++ обменника и нарушение внутриклеточной мобилизации кальция, вероятно, опосредованное цАМФ, могут быть важными компонентами описанного спазмолитического действия липидного экстракта [118].
A. Tamalunas и соавт. продемонстрировали ингибирующее действие HSRE на сокращение гладких мышц предстательной железы и мочевого пузыря человека. Применение HSRE вызывало дозозависимое ингибирование сокращения гладкомышечных элементов предстательной железы, вызванное адренергической стимуляций, на 30–60%, что подтвердило неконкурентное связывание компонентов экстракта с α1-адренорецепторами [120, 121]. Это дополнило и дало новое объяснение ранее выявленному благоприятному эффекту SRE, полученных разными методами экстракции, в отношении широкого спектра агонистов сокращения в богатых гладкими мышцами органах, в том числе в нижних мочевыводящих путях, что указывает на неконкурентное связывание с α1-адренорецепторами как на классовый эффект SRE, при том, что активность и эффективность экстрактов различались [110, 112].
SRE, помимо действия на адрено – и холинорецепторы мочевого пузыря, потенциально способен ингибировать ваниллоидные рецепторы 1-го типа (TRPV1). Эти рецепторы локализуются в эпителии мочевого пузыря, участвуют в нейрогенной регуляции мочеиспускания, вероятно, во взаимодействии с TRPV1 афферентных нервных волокон и способствуют передаче ощущения желания опорожниться в мозг. Активацию TRPV1 отмечали при развитии гиперактивности детрузора. Приведенные факты объясняют эффективность SRE в отношении симптомов фазы накопления у больных ДГПЖ [122].
Хотя считают, что основными компонентами, обуславливающими терапевтическую эффективности SRE, являются липофильные соединения, которые в большей степени должны накапливаться в железистых органах (предстательная железа), чем в мышечных органах (мочевой пузырь), накопление активных компонентов в мочевом пузыре в реальных концентрациях также происходит [112, 114, 116]. При этом накопление агента в ткани необязательно приравнивается к его специфической активности в этой ткани, если активность агента связана с накоплением в специфичных мембранах, а активность обусловлена локализацией внутри мембраны. Этот тезис подтверждается рядом исследований, показавших, что пероральное введение SRE животным значительно ослабляло уродинамические симптомы при гиперактивном мочевом пузыре не только у самцов, но и у самок крыс за счет увеличения емкости мочевого пузыря и интервалов между мочеиспусканиями, что подтверждает прямое влияние препарата на мочевой пузырь [119, 123].
В совокупности приведенные результаты свидетельствуют о том, что эффективность SRE в отношении СНМП также обусловлена его способностью регулировать активность фармакологически значимых рецепторов в мочевом пузыре: α1-адренорецепторов; М-холинорецепторов и ваниллоидных рецепторов.
ЗАКЛЮЧЕНИЕ
Как было показано выше, патогенез ИВО тесно связан с механическим напряжением, гипоксией стенки мочевого пузыря, воспалением и окислительным стрессом, что подразумевает комплексную реакцию организма, реализуемую через множество факторов, включая критическую роль системного иммунного ответа. Анализ изложенных в литературе результатов экспериментальных и клинических исследований свидетельствует, что на уровне ПЖ HSRE демонстрирует комплексный механизм антиандрогенного, антипролиферативного и противовоспалительного действия. Наше недавнее исследование [6] и ряд других, описанных выше, указывают на возможность влияния HSRE и на мочевой пузырь.
И, наконец, S. Yamada и соавт. в недавнем клиническом плацебо-контролируемом исследовании продемонстрировали статистически значимое сокращение числа дневных и ночных мочеиспусканий при лечении SRE гиперактивного мочевого пузыря у женщин, что, безусловно, свидетельствует о влиянии SRE на функцию мочевого пузыря [124].
Однако механизмы такого действия не ясны. Рассмотрение основных путей патогенеза ИВО, в сопоставлении с известными данными о влиянии SRE на ПЖ и отдельными исследованиями о воздействии SRE на мочевой пузырь, теоретически дает понимание точек возможного приложения экстракта Serenoa repens при дисфункции мочевого пузыря. Концептуально, механизмы позитивного плейотропного влияния HSRE на обструктивный мочевой пузырь могут быть представлены следующей схемой (рис. 4).
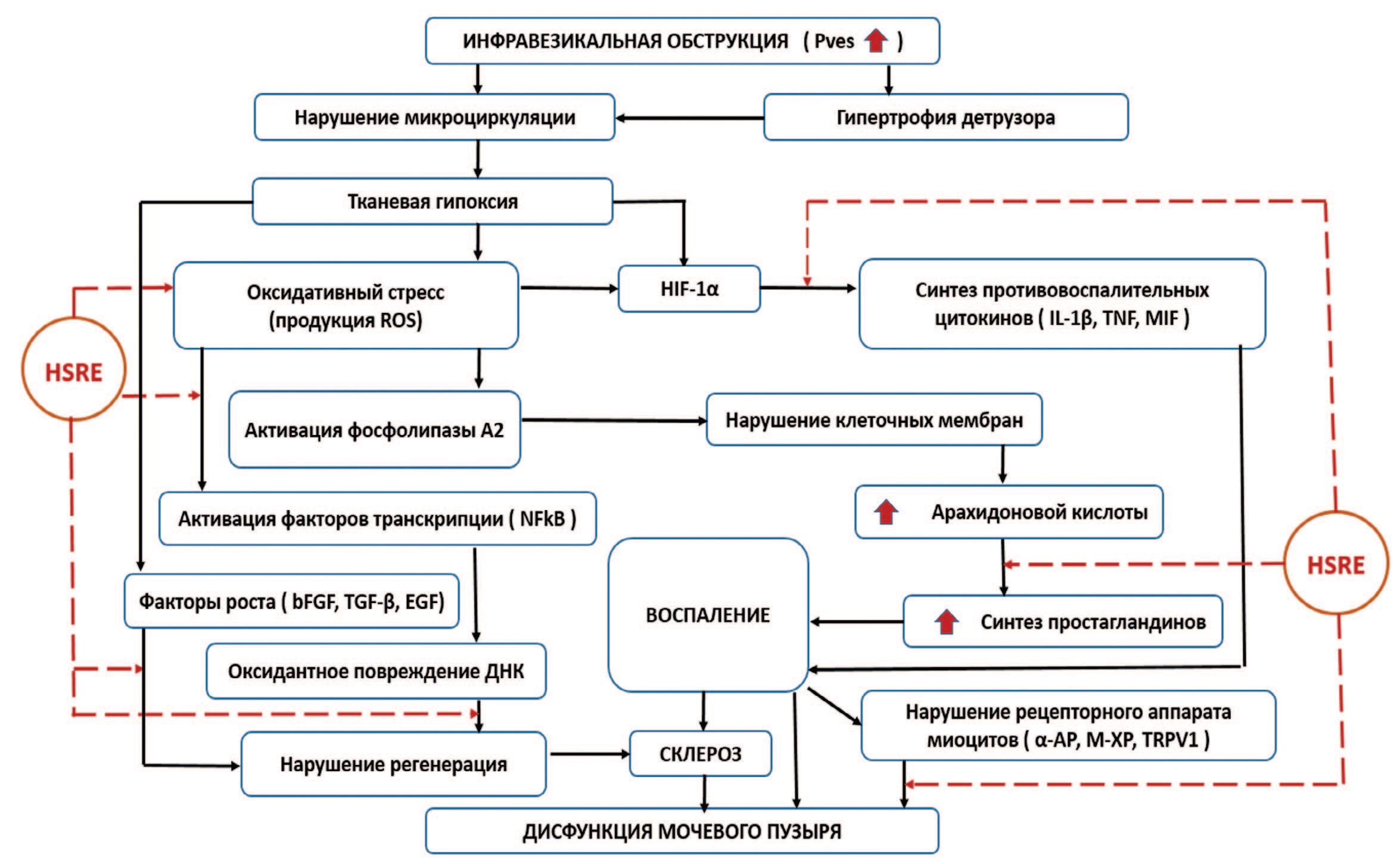
Рис 4. Схема возможного влияния HSRE на звенья патогенеза дисфункции мочевого пузыря при ИВО
Fig. 4. Scheme of possible influence of HSRE on the links of the pathogenesis of bladder dysfunction in IVO
Основные звенья этого воздействия – следующие:
- торможение развития патологического процесса на этапе гипертрофии – начале фазы декомпенсации;
- протективное действие HSRE путем антиоксидантной активности;
- противовоспалительный эффект, проявляющийся снижением IL-1β, включением защитных механизмов, связанных с влиянием на сигнальные пути HIF-1, NF-κB и MIF;
- регуляция каскада арахидоновой кислоты и снижение синтеза простагландинов (противовоспалительное действие);
- регуляция факторов роста, в первую очередь bFGF, TGF-β и EGF;
- воздействие на регуляцию α1-адренорецепторов, М-холинорецепторов и ваниллоидных рецепторов.
Тем не менее, при анализе экспериментальных работ следует учитывать, что человеческий детрузор значительно отличается от моделей на животных, используемых для исследований, которые не страдают от естественной ИВО [125]. Однако общие биологические механизмы имеют место быть, а, вероятно, неспецифическое действие экстрактов Serenoa Repens, подтвержденное на предстательной железе, должно проявляться и на мочевом пузыре.
